![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
|
味のお話
味覚は、ホメオスタシスを維持する生理機能の一環を担う。例えば体内の糖、必須アミノ酸や電解 質が欠乏すると、生物はその物質の「味覚」を手掛かりとして欠乏物質を探り出し摂取する。逆に、過剰になると摂取するのをやめる。これらの食行動の変化は、おいしい、まずいといった快不快の情動、咀嚼運動、唾液、消化液、ホルモン、満腹物質の分泌など、食にかかわる神経系や液性の様々な調節系が働くことによりもたらされる。動物の生息域が変われば、食環境も変わる。生物は系統発生や進化の過程で、味覚感受性やその支配遺伝子がその長い年月をかけた営みの中で、食環境に適応すべく変容を遂げてきたものと思われる。事実、食性の異なる動物種間では味覚感受性に顕著な差が認められるし、ヒトの個体間人種間でも味物質に対する感受性は大きく異なっている。したがって、生物の味覚感受性の多様性とその表現型を明らかにすることは、味覚センサーの機能を明らかにするだけでなく、食環境にあわせたその分子進化(モーダルシフト)の過程をも理解することになると思われる。本稿では、近年ようやくその謎がとかれつつある味覚センサー(特に脊椎動物の甘味/苦味受容体)の機能、その多様性と表現型との連関について、最近の知見を紹介する (細胞工学; 2007: 890-3より改編引用)。
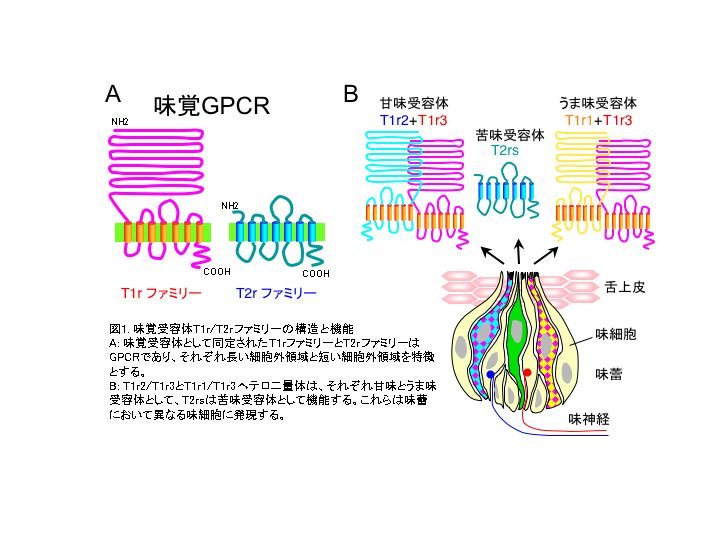
I. 味覚受容の謎の解明: 味覚センサー分子T1r, T2rの発見 脊椎動物の味覚は、口腔内の味覚受容細胞により知覚される。この細胞は様々な化学物質に応答し、その情報を中枢に伝える。味覚は基本的に5つの味質(甘味、塩味、酸味、苦味、うま味)に分類される。 これらの中で、甘味(カロリー源)、塩味(ミネラル)、うま味(タンパク質)は、好ましい味で、その物質を体内へ摂取する方向に作用し、一方、酸味や苦味は嫌な味で、腐敗物や毒の摂取を防ぐシグナルとなる。甘味、うま味、苦味受容体については、2つのGPCRファミリー(T1rとT2rファミリー)が同定されたことにより、味覚受容の謎が急速に解き明かされつつある1)。 マウスの甘味感受性は、第4染色体上の2つの遺伝子座(dpa (D-フェニルアラニン(D-Phe)甘味感受性)とその遠心位のSac(サッカリン嗜好性)によって支配されていることが感受性の系統差を利用した遺伝学的解析により推測されていたが、長年その実体については不明であった。しかし2001年、ゲノムデータベース解析により、そのうちの1つのSac遺伝子座及びその近傍にT1r1, T1r2, T1r3(Sac 遺伝子支配)が同定された。これら3つのメンバーはT1rファミリーと呼ばれ、GPCR クラスCファミリー(mGluRs、Ca受容体、V2rフェロモン受容体)に属し、複数のイントロンと長い細胞外領域を特徴とする。味細胞では、T1r3はT1r1もしくはT1r2と共発現する。HEK細胞強制発現系を用いた解析では、T1rsは単独では応答せず、T1r3をT1r2もしくはT1r1と共発現させた場合にのみ、それぞれ甘味、うま味物質に応答することが示された。さらにT1r1、T1r2、T1r3遺伝子欠損マウスでは、それぞれうま味、甘味、うま味と甘味の両方の感受性がなくなったことにより、T1r2/T1r3ヘテロ二量体が甘味の受容体として、T1r1/T1r3がうま味の受容体として機能していることが明らかにされた。(図1)このT1r2/T1r3の機能的特徴は、化学構造の異なる様々な甘味物質(天然糖、人工甘味料、甘味アミノ酸、甘味タンパク質)を受容する、いわゆる低リガンド特異性である。 マウスの苦味感受性に関しては、第6染色体の遠位に複数の苦味感受性支配遺伝子[sucrose octaacetate (SOA)、cycroheximide (Cyx)、quinine (Qui)、raffinose acetate (Rua)、copper glycinate (Glb)]の存在が示唆されていた。T2rファミリーは、T1rsと同様に、この染色体上に存在するGPCRをゲノムデータベース解析することにより同定された。T2rsはGPCR クラスAファミリー(V1rフェロモン受容体やopsin受容体)に属し、イントロンを含まず、短い細胞外領域が特徴である。また味細胞において、T2rsはT1rsと共発現しないことが明らかにされている(図1)。これまでにヒトでは少なくとも26種類、マウスとラットでは35種類が明らかにされており、マウスT2r5 (mT2r5)とラットT2r9 (rT2r9)はcycloheximide、mT2r8とヒトT2r4 (hT2r4)はdenatonium、hT2r16はsalicin、hT2r10はstrychine、hT2r14はpicrotoxinin、T2r38は phenylthiocarbamide(PTC)と6-n-propylthiouracil(PROP)を特異的に受容することが報告されている2)。
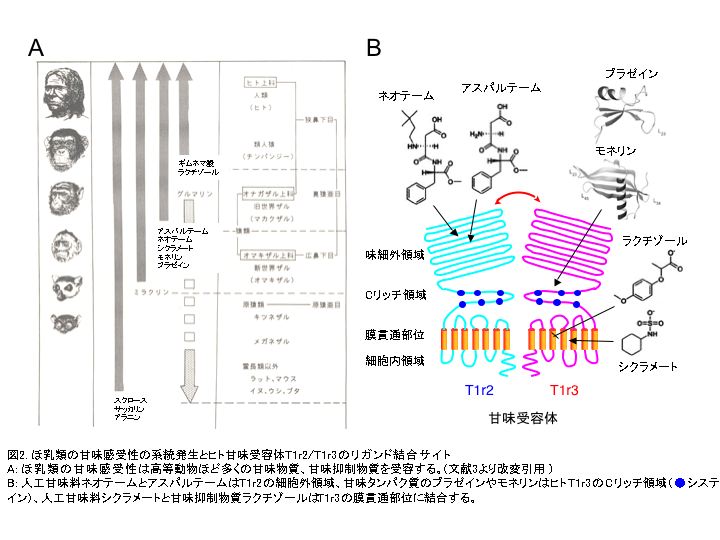
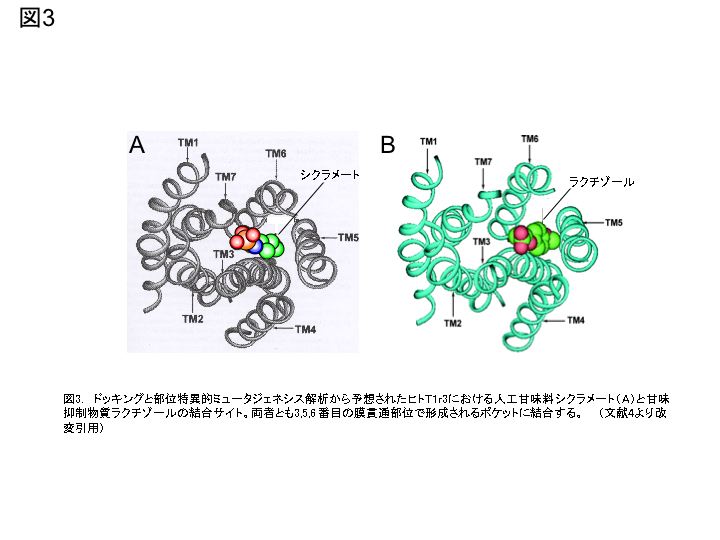
甘味受容体は系統発生的に進化しており、高等動物ほど多くの物質を受容すると考えられている(図2A)。ショ糖やブドウ糖などの糖、グリシンやアラニンなどの甘味アミノ酸はほとんど全てのほ乳動物に甘味をもたらす。しかし、熱帯植物の果実から抽出した、酸味を甘酸味に変える性質のミラクリンは、霊長類以外のほ乳動物や霊長類でも最も下位に位置するロリスなどの原猿類(原猿亜目)には無効であり、リスザルなど新世界ザル以上のいわゆる真猿亜目に属する動物で有効となる。また、植物由来の高甘味タンパクのモネリンやタウマチン、人工高甘味ペプチドのアスパルテームやネオテームは、真猿亜目の中でも新世界ザル(広鼻下目)以下の動物は応答せず、旧世界ザル以上(狭鼻下目)の動物のみが応答する。さらに、甘味抑制物質のギムネマ酸は、狭鼻下目でもマカクザルなどオナガザル上科には無効で、ヒトやチンパンジーなどヒト上科ではじめてその甘味抑制効果が出現する。奇妙なことに、同じ植物(ギムネマシルベスタ)から抽出されたグルマリンはマウス、ラットおよびマカクザルの甘味応答を一部抑制するが、ヒトの甘味は抑制しない3)。これらの甘味受容の顕著な差の謎が、T1rsのリガンド結合サイトの解析により解き明かされつつある。 T1rsの構造的特徴は、サブファミリー分子であるmGluR1のX線結晶構造解析により明らかにされている。7回膜貫通領域に続くシステインを多く含む (C-rich) 領域を含む大きな細胞外領域、そして下流のGタンパク質を活性化させる細胞内領域を有し、ホモあるいはヘテロ二量体を形成して機能するという特徴をもつ。細胞外領域の約2/3はローブ1とローブ2という構造的ユニットを形成するが、両者の構成する境界面が開閉することから“Venus flytrap”と呼ばれ、リガンドの認識と結合に関与することが知られている。 T1r2/T1r3におけるリガンド結合サイトは、感受性が顕著に異なるヒトとマウスT1r2/T1r3の細胞外領域や膜貫通部位を様々なパターンで組み換えたミュータントを、HEK細胞に強制発現させ、その応答特性を調べることで明らかにされた。これまでに人工甘味料であるアスパルテームとネオテームの受容にはヒトT1r2のN末端のVenus flytrap領域、甘味タンパク質であるブラゼインにはヒトT1r3のC-rich領域、人工甘味料シクラメートと甘味抑制物質ラクチゾールにはヒトT1r3の膜貫通領域がそれぞれ結合に重要であることが報告された(図2B)。さらに近年、ドッキングと部位特異的ミュータントの解析から、アスパルテームは、主にT1r2のAsp278/Asp307(塩橋)、Ser303/Arg383/Val384(水素結合)とTyr215/Tyr103/Pro277(疎水相互作用)に結合し、ブラゼインは、T1r3のリガンド受容に関わるC-rich領域(6つのシステインが3つのS-S結合を形成)のAla537とPhe540の変異により応答がみられなくなること、シクラメートは、T1r3の3, 5, 6番目の膜貫通部位のポケットに、またラクチゾールの結合に関わるT1r3のHis641/Arg723(塩橋と水素結合)とPhe778/Leu782/Leu644(疎水相互作用)はシクラメートの結合サイトと重なることが明らかにされている(図3)。このように、T1r2/T1r3の低リガンド特異性や感受性の多様性は、両遺伝子にある複数のリガンド結合サイトの存在とそれらの変異によることが明らかになってきた4)。
味覚感受性の多様性と味覚受容体の遺伝子多型性との連関も明らかになってきた。マウスにおいて、mT2r5がシクロヘキシミドをリガンドとすることは先に述べた。このことが明らかにされたのは、感受系統のDBA/2Jと非感受系統のC57BL/6JのT2r5間に見いだされた5つのアミノ酸変異を伴うSNP (1塩基変異)が味覚感受性低下の原因であることが明らかにされたためである2)。甘味感受性のマウス系統差においては、T1r3の60番目のアミノ酸変異(Ile60The)が、その原因の1つであることが示唆されている1)。また、ネコは甘味を感じないことが知られているが、T1r2のコーディング部分に247bpの遺伝子欠損があり、機能的T1r2が発現しないことが報告されている5)。 ヒト味覚感受性の個人差についての最初の報告は、Fox (1931年)によるPTC(N-C=S基を持つ)に対するもので、“taste blindness: 味盲”として紹介された。味覚閾値分布は2峰性を示し、感受性群と非感受性群では約1000倍の濃度差がみられる。近年、人種差を利用した遺伝学的解析により、第7染色体上のT2r38がその原因遺伝子として同定された。この背景となったのは3ヶ所のアミノ酸変異( pro49→Ala、Ala262→Val、Val26→Ile)がその鍵となり、ハプロタイプPAV(pro49/Ala262/Val26)型は感受性、AVIは非感受性、またAAV, AAI, PVIはそれらの中間の表現型を示すことが明らかにされたためである。表現型の地域分布についても報告されており、西洋、アジアとアフリカでは、AVIとPAVが最も多くみられ(西洋とアジアでは他のタイプはみられなかったが、アフリカでは AAIもみられる)、ネイティブのアメリカ人はPAVのみであることが明らかとなった2)。ヒトと最も近いボノボとチンパンジーの3種間でT2rを比較した報告では、ファミリーの構成はほとんど同じであったが(特にヒトとボノボでは98%以上)、種特異的に偽遺伝子化(bT2r9、hT2r62、chT2r64、bhT2r63)や、複数の遺伝子多型がみられることが明らかとなった6)。これらのことから、ヒトにおいても遺伝子多型性とその表現型との間には直接的な関係があること、それは生活環境(この場合、苦味物質の地域分布)と強くリンクし、さらに人類の祖先が6-7Maに中央アフリカに誕生してから、今もなおその変化が続いていることが明らかになってきた。
味細胞で受容された味覚情報は味神経を介して中枢へ伝えられます。その投射経路はラットやサルでよく調べられています。ラットでは味覚神経はまず延髄の孤束核に入ります。孤束核は吻尾的に細長い構造をしており、吻側から尾側にわたって舌前方の味覚情報を伝える鼓索神経、舌後方の舌咽神経、喉頭部の上喉頭神経が順に終止します。孤束核からの味覚性二次ニューロンは大きく2つに分かれます。一つは味覚にもとづく顔面表情変化や唾液、消化液、インスリン分泌といった体性運動、消化、血液循環、呼吸、内分泌など反射性活動に関与する経路で、毛様体に入ったあと、三叉神経運動核、顔面神経核、舌下神経核、迷走神経背側運動核、疑核などにいきます。もう一つの経路は次の味覚中継核である橋の結合腕傍核へ同側性に入り、そこから視床味覚野、そして皮質味覚野へと投射されます(背側路)。また、結合腕傍核からは視床下部、不碓帯、扁桃体への投射もみられます(腹側路)。背側路は味の強さや質の分析をおこない、腹側路は味の快や不快の決定や食欲に関係するものと考えられています。サルでは基本的にラットと同じ経路であると考えられていますが、結合腕傍核は経由しないと考えられています。 最後に、それぞれの味はすべて味神経の電気信号に変換され、その信号のしくみ自体は共通です。 おわりに げっ歯類やヒトなどの味覚感受性の「種差」「系統差」「個体差」を利用した解析を行うことにより、T1r/T2r味覚受容体の機能、構造やリガンド特性のみならず、環境による味覚受容体の進化の過程までもが少しずつ見えはじめてきた。ヒトについては、今後さらに遺伝子多型性について解析することにより、味覚感受性と食環境や健康との相関性がより鮮明に浮き上がってくるものと思われる。現在、我々の研究室ではヒト味覚感受性とT1rs遺伝子多型性との相関解析を進めているが、世界でもっとも繊細な舌をもつといわれる?日本人特異的な遺伝子型が明らかになるかもしれない。将来的には、これらのデータをもとにして、摂食と直接リンクする生活習慣病(肥満、糖尿病、高血圧など)の予防や治療に応用できるより効果的な甘味・塩味修飾(カロリー・ナトリウム調節)物質や、健康食品や医薬品の苦味抑制物質などが開発され、さらには個人の味覚・遺伝情報をもとにした疾病予測ツール、舌切除などによる味覚喪失者への人工味覚器の開発なども現実のものとなるかもしれない。 引用文献: 1) Chandrashekar J, et al: Nature (2006) 444:288-294 2) Meyerhof W: Rev Physiol Biochem Pharmacol. (2005) 154: 37-72 3) 二ノ宮裕三: 味覚感受性と遺伝(佐藤昌康・小川尚編、最新味覚の科学、朝倉書店、東京)37-47, 1999 4) Cui M, et al: Curr Pharm Des. (2006) 12: 4591-4600 5) Li X, et al: J Nutr. (2006) 136:1932S-1934S 6) Parry CM, et al: Proc Natl Acad Sci U S A. (2004) 101:14830-14834 重村憲徳、大栗弾宏、實松敬介、二ノ宮裕三:味覚センサーの分子進化と多様性・細胞工学; 2007: 890-3 二ノ宮裕三、重村憲徳:基本解説1:p63-70・味のなんでも小事典:甘いものはなぜ別腹?・日本味と匂学会編・ブルーバックスシリーズ・講談社 |